
Engineered mammalian and bacterial extracellular vesicles as promising nanocarriers for targeted therapy

Abstract
Extracellular vesicles (EVs), which are nanocarriers with phospholipid bilayer structures released by most cells, play a key role in regulating physiological and pathological processes. EVs have been investigated due to their loading capacity, low toxicity, immunogenicity, and biofunctions. Although EVs have shown good potential as therapeutic vehicles, natural EVs have a poor targeting ability, which substantially reduces the therapeutic effect. Through the addition of a targeting unit into the membrane surface of EVs or inside EVs by engineering technology, the therapeutic agent can accumulate in specific cells and tissues. Here, we focus on mammalian EVs (MEVs) and bacterial EVs (BEVs), which are the two most common types of EVs in the biomedical field. In this review, we describe engineered MEVs and BEVs as promising nanocarriers for targeted therapy and summarize the biogenesis, isolation, and characterization of MEVs and BEVs. We then describe engineering techniques for enhancement of the targeting ability of EVs. Moreover, we focus on the applications of engineered MEVs and BEVs in targeted therapy, including the treatment of cancer and brain and bone disease. We believe that this review will help improve the understanding of engineered MEVs and BEVs, thereby promoting their application and clinical translation.
Keywords
INTRODUCTION
According to the International Society for Extracellular Vesicles, extracellular vesicles (EVs) are defined as “particles naturally released from the cell that is delimited by a lipid bilayer and cannot replicate”[1]. Notably, the term “exosomes” is often used as a general description of EVs[2,3]. EVs can transmit a variety of biologically active components, such as proteins, nucleic acids, lipids, and metabolites, to affect the performance of recipient cells[4-7]. Many studies have found that EVs play an important role in regulating the physiological and pathological processes of the body by participating in cell-to-cell communication, cell proliferation, cell migration, angiogenesis, and immune regulation[6,8]. Due to their unique nanosized structures, loading capacity, biochemical properties, and good biocompatibility, EVs have been widely used in various applications in the biomedical field, such as vaccines, cancer agents, and drug delivery vehicles[9-11]. Although EVs have shown good potential as therapeutic vehicles, natural EVs were shown to have a poor targeting ability in animal experiments, which substantially reduced the therapeutic effect[12,13]. Therefore, many engineered methods have been applied to improve the targeting ability of EVs[14-16].
In our previous study, we focused on engineering EVs derived from mammalian cells such as endothelial cells[17] and NIH-3T3 cells[18]. Although these engineered mammalian EVs (MEVs) were shown to have an excellent targeting ability and therapeutic effect, the low extraction yield (requiring many mammalian cells) is still a limiting factor. The current complex and low yield protocols for purification and extraction of EVs, such as ultracentrifugation, gradient ultracentrifugation, co-precipitation, size-exclusion chromatography, and field flow fractionation, pose a tremendous challenge in the mass production of EVs. Therefore, we recently paid more attention to bacterial EVs (BEVs), which can be easily obtained through fed-batch fermentation and purification procedures[4,13]. Moreover, according to the latest minimal information for studies of extracellular vesicles (MISEV) in 2021, the topic of “nonmammalian EVs, especially BEVs” ranked fourth[1]. Although engineered BEVs have also been used in the field of biomedicine[19,20], BEV research is less developed than that of MEVs. The number of MEV and BEV studies has increased rapidly in recent years (PubMed.gov). In general, MEVs and BEVs are the two most common types of EVs in the biomedical field.
Due to the importance of MEVs and BEVs in the field of biomedicine, we focus on these two types of EVs and their engineering and applications in this review. Here, to elucidate engineered MEVs and BEVs as promising nanocarriers for targeted therapy, we first summarize the biological basis of MEVs and BEVs, including different mechanisms of biogenesis, isolation, and characterization. We then present approaches for modifying BEVs and MEVs, which are physical engineering (membrane fusion and membrane coating), biological engineering [membrane fusion, lysosome-associated membrane glycoprotein 2B (LAMP-2B), and CD63], and chemical engineering (covalent reaction and noncovalent reaction), to enhance the targeting ability. Finally, we conclude with the application of engineered MEVs and BEVs in targeted therapy of tumors (chemotherapy, gene therapy, photothermal therapy, and immunotherapy), brain disease [Alzheimer’s disease (AD), Parkinson’s disease (PD), and ischemic stroke], and bone disease [osteoarthritis (OA) and osteoporosis (OP)]. This review will help improve our understanding of the importance of MEVs and BEVs and thus promote targeted therapy for various diseases.
THE BIOGENESIS OF MEVS AND BEVS
EVs are a general term for nanovesicles with phospholipid bilayer structures secreted by most cells[2,3,21]. EVs can be secreted by almost all cells and are widely present in cell supernatants and various body fluids[22]. As early as the 1960s, BEVs were first reported in the Gram-negative bacteria Escherichia coli[23-26]. In the 1980s, Pan and Harding et al.[27,28]successively observed the release of MEVs in reticulocytes. At this stage, both BEVs and MEVs were regarded as “garbage bags” for cells to discharge metabolic waste[29]. In 1996, Raposo et al.[30] found that EVs derived from B lymphocytes can present antigens and activate T lymphocytes to participate in the regulation of immune cells. Soon after, EVs that could transfer nucleic acids such as mRNA and miRNA were also found in archaea[31,32]. Gradually, researchers discovered that the role of EVs is much more than clearance of cell waste; EVs also transmit signals to distant parts of the body, where they can affect multiple dimensions of cell life[10]. A detailed description of the mechanisms of MEVs and BEVs would provide an important theoretical basis for the treatment of disease.
The biogenesis of MEVs
According to the size, biological characteristics, and formation process, MEVs can be classified into three major subtypes: exosomes, microvesicles, and apoptotic bodies [Figure 1A][33]. Exosomes are EVs with a diameter of 40-160 nm formed by the fusion of multivesicular bodies (MVBs) and cell membranes. Moreover, microvesicles are EVs with a size range of 200-1000 nm in diameter that are directly formed by cell membrane budding. Apoptotic bodies are vesicular bodies with larger diameters (500-2000 nm in diameter) formed by cell fragmentation during the process of cell apoptosis. Among these subtypes, exosomes have received widespread attention due to their sizes, biological composition, and cell-to-cell communication ability[10]. Therefore, we use exosomes to represent MEVs in this review.
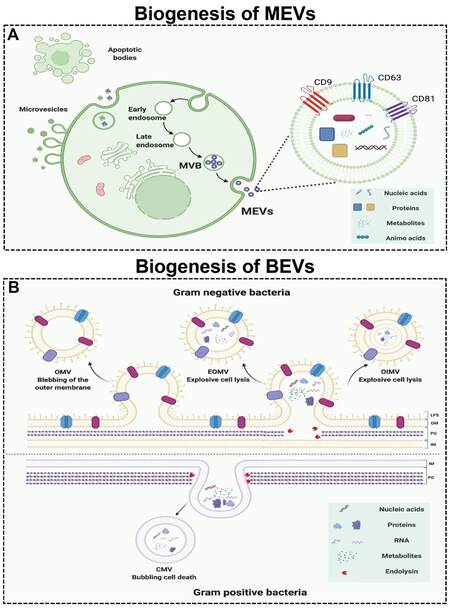
Figure 1. The biogenesis of mammalian extracellular vesicles (MEVs) and bacterial extracellular vesicles (BEVs). (A) The biogenesis of MEVs. (B) The biogenesis of BEVs. Figures were created with Biorender.com.
MEVs are formed by the endosomal system in a process involving three stages [Figure 1A][34]. First, the plasma membrane invaginates to form endocytic vesicles, which fuse with each other to form early endosomes. Second, early endosomes invaginate again to encapsulate intracellular cargos, forming multiple intraluminal vesicles (ILVs), which are further transformed into late endosomes, MVBs. Finally, the MVBs fuse with the plasma membrane and excrete their contents into the extracellular space. The formation, sorting of cargos, and release of exosomes are a series of finely regulated processes that require the participation of many proteins. The formation of exosomes involves proteins such as endosomal sorting complex required for transport (ESCRT), transmembrane proteins (CD9, CD63, and CD81), apoptosis-linked gene 2-interacting protein X (Alix), and tumor susceptibility gene 101 protein (TSG101). Moreover, the intracellular transport of exosomes involves the participation of many molecular switches such as the RAB GTPase protein and cytoskeletal proteins such as actin and tubulin[35]. In addition, the secretion of exosomes requires the participation of SNARE protein complexes and the synaptic binding protein family[36]. The protein composition of exosomes can reflect the characteristics of their endosomal origin. In different types of cells and body fluids, exosomes all contain the same marker protein molecules, such as Alix, TSG101, SNARE, and RAB GTPase, and the transmembrane proteins, CD9, CD63, and CD81[37]. In addition to specific protein composition, exosomes also have a special lipid composition. Exosome membranes are enriched with cholesterol, ceramide, and sphingolipids[38]. These lipids are also involved in the formation and secretion of exosomes. For example, ceramide is involved in the budding of ILVs and MVBs[38].
The biogenesis of BEVs
Bacteria are divided into Gram-negative (G-) bacteria and Gram-positive (G+) bacteria based on their structure, morphology, and staining properties. Both G+ and G- bacteria can release EVs without energy consumption[39,40]. BEVs are EVs with sizes of 20-400 nm in diameter and can be divided into four types: outer-membrane vesicles (OMVs), explosive outer-membrane vesicles (EOMVs), outer-inner membrane vesicles (OIMVs), and cytoplasmic membrane vesicles (CMVs). The first three EVs are formed by G- bacteria, and the latter are formed by G+ bacteria. G+ bacteria produce CMVs by endolysin-triggered cell lysis (bubbling cell death) [Figure 1B][26]. OMVs are formed by blebbing of the outer membrane of G- bacteria, while OIMVs and EOMVs are released by explosive cell lysis of G- bacteria [Figure 1B][25].
Similar to MEVs, BEVs are lipid bilayer-enclosed structures containing various biomolecules released by cells and are increasingly regarded as the main form of cell-to-cell communication[41-43]. Due to the diversity of their contents, BEVs have a key role in bacteria-bacteria and bacteria-host communications. Generally, BEVs contain high levels of proteins, nucleic acids, metabolites, small molecules, etc. G- BEVs are enriched in periplasmic proteins such as the multidrug efflux pump subunit AcrA and outer membrane proteins such as outer membrane protein F (OmpF). Notably, Vanaja et al.[44] used OmpF as a specific (surface) marker for E. coli-derived EVs. However, the lack of specific markers is still a challenge in the field of BEVs[39]. Moreover, lipids are an important structural component of bacterial cell membranes. The most significant difference between G+ and G- BEV contents is lipopolysaccharide (LPS, or endotoxin), which can cause an innate immune response[45]. Knockout of msbB in G- bacteria E. coli for less endotoxic EVs is a common approach[15,19,20,46]. Interestingly, the nonpathogenic G- bacteria E. coli Nissle 1917 lacks definite virulence factors such as LPS, so it can be used as a probiotic for the treatment of various gastrointestinal diseases[47-49]. Due to its easy genetic manipulation and probiotic characteristics, E. coli Nissle 1917 and its BEVs are promising candidates for medical engineering. In addition, BEVs have been reported to transfer nucleic acids, such as DNA and RNA, into other bacterial cells[50] and mammalian cells, which trigger different host immune responses and cellular processes[51,52]. RNA, especially miRNA and siRNA, can be protected from degradation through BEVs, which promotes delivery to mammalian cells[53]. BEVs selectively package different metabolites depending on the strains. Gujrati et al.[15] reported that the BEVs secreted by strains overexpressing melanin (Mel) also contain Mel. These findings indicate the applications of EVs in biomedicine.
THE ISOLATION OF MEVS AND BEVS
The isolation and characterization of MEVs and BEVs is an indispensable step for their further application in biomedicine. In fact, the isolation of such nanoparticles is generally difficult. MEVs can be derived from a variety of biological fluids, such as blood serum[54], breast milk[55], urine[56], tears[57], saliva[58], and sperm[59]. However, BEVs are found in many kinds of media, such as LB, MRS broth[60], and BHI broth (which always requires porcine mucin for Akkermansia muciniphila)[61]. Therefore, there is a major difference in viscosity, which causes difficulties in isolation and purification. Moreover, the amount of sample available for isolation is another factor that affects efficiency. Recently, various isolation methods, such as differential centrifugation, precipitation, size exclusion chromatography, and magnetic capture, have been established based on the differences in size, density, charge, and surface ligands[62-65]. Here, we summarize the most commonly used and effective MEV and BEV isolation techniques [Figure 2].
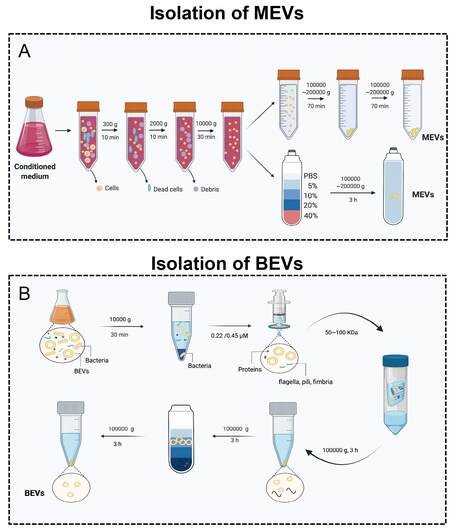
Figure 2. The isolation of MEVs and BEVs. (A) The isolation of MEVs. (B) The isolation of BEVs. Figures were created with Biorender.com.
Isolation of MEVs
Ultracentrifugation-based MEV isolation is the gold standard, thus one of the most commonly used and reported techniques[66-69]. According to the first large-scale detailed survey of current global MEV isolation practices, 81% of researchers used ultracentrifugation (including differential centrifugation) for MEVs isolation[68]. Li et al.[67] summarized the use of ultracentrifugation and differential centrifugation to remove other impurities in the sample through a combination of different speeds and times to finally achieve the isolation of MEVs. Low-speed centrifugation (300-2000 g or 2000-10,000 g) is generally used to remove cells, dead cells, cell debris, etc. Ultracentrifugation (100,000-200,000 g) is generally applied to collect MEVs [Figure 2B]. In addition, for better purification of MEVs, density gradient centrifugation such as iodixanol can be used [Figure 2A]. After the isolation of MEVs, the most commonly used characterization methods are transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA), and Western blotting (WB)[17,18,70,71]. TEM and NTA are used to show the sizes, shapes, and concentrations of EVs. The transmembrane proteins CD9, CD63, and CD81, as well as TSG101, are often used as specific markers in WB[17,18,70,71].
Isolation of BEVs
The standard protocol for BEVs purification is to physically separate EVs from cell culture through a series of steps [Figure 2B][72-74]. Simply, low-speed centrifugation (2000-10,000 g) is used to remove bacteria and their debris in the fermentation broth. Then, a 0.22 μm sterile filter is applied to remove residual bacteria. Subsequently, a 100 kDa ultrafiltration membrane is required to remove non-BEV-associated proteins. Furthermore, ultracentrifugation and density gradient centrifugation are used together for the separation and purification of BEVs. Using the above method, we successfully obtained multiple BEVs, such as EVs derived from Lactobacillus rhamnosus GG[4]. Similar to the characterization of MEVs, TEM and NTA are common methods used to assess BEVs. The outer membrane proteins OmpA[20] and OmpF[44] are used as specific markers for E. coli-derived EVs. However, many studies involving BEVs do not use WB[15,19,75]. The selection of specific markers is still a major challenge in the BEV field.
ENGINEERING TECHNIQUES TO IMPROVE THE TARGETING ABILITY OF MEVS AND BEVS
Nanosized EVs have been investigated as therapeutic vehicles due to their loading capacity, low toxicity, immunogenicity, and biofunctions[10,76]. However, the poor targeting ability of natural EVs is not conducive to therapy. With the introduction of the concept of precision medicine in 2015[77], researchers are increasingly investigating the targeting ability of EVs. Targeted delivery could increase the local concentration of the therapeutic agent and minimize side effects. Through the addition of a targeting unit into the membrane surface of EVs or inside EVs by engineering technology, the therapeutic agent could accumulate in specific cells and tissues. Many engineering technologies such as biological engineering and chemical modification have been used to modify EVs to enhance their targeting ability. The membrane surface of MEVs is rich in lipoproteins (such as phosphatidylserine, cholesterol, sphingomyelin, and ceramide) and membrane proteins (adhesion molecules, integrins, membrane transport proteins, MHC class I/II, tetraspanins, and transferrin receptor)[78]. At present, the targeting engineering of MEVs is intensively studied[13,16,66], but research on the targeting engineering of BEVs has just started[15,20]. Their similar phospholipid bilayer structure makes most engineering methods universal. Here, we summarize the techniques to improve the targeting ability of MEVs and BEVs [Figure 3 and Table 1].
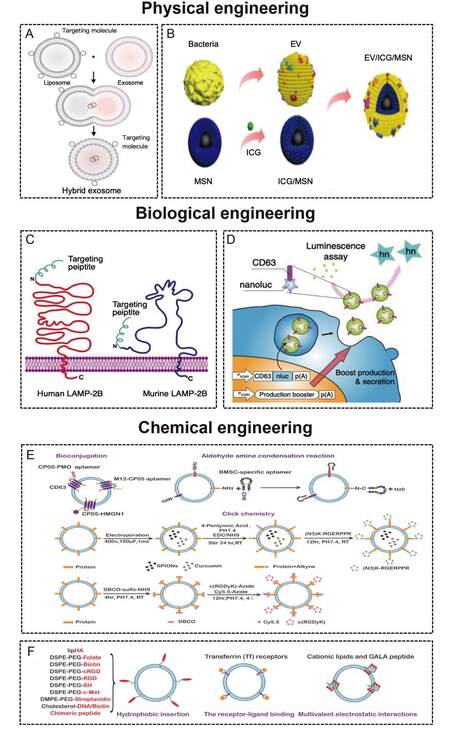
Figure 3. (A) The procedure to produce hybrid nanoparticles by membrane fusion. Liposomes with targeting molecules on the surface can be delivered into EVs through membrane fusion[79]. Copyright 2018 WILEY-VCH. (B) The procedure to produce hybrid nanoparticles by membrane coating[80]. Copyright 2020 Ivyspring International Publisher. (C) The fusion of the targeting peptide with LAMP-2B[66]. Copyright 2021 Ivyspring International Publisher. (D) The fusion of the delivery molecule with CD63[81]. Copyright 2018 Springer Nature. (E) Targeted modification of EVs based on chemical covalent reactions[82]. Copyright 2021 Elsevier. (F) Targeted modification of EVs based on chemical non-covalent reactions[82]. Copyright 2021 Elsevier. LAMP-2B: Lysosome-associated membrane glycoprotein 2B.
Summary of the techniques to improve the targeting ability of MEVs and BEVs
| Methods | Target cells/Tissue | Ref. |
| Physical engineering | ||
| Membrane fusion | Mesenchymal stem cells Bone mesenchymal stem cells | [79] [18] |
| Membrane coating | Lymph nodes | [80] |
| Biological engineering | ||
| LAMP-2B | Neuronal cell Colorectal cancer (HCT-116) Synovial mesenchymal stem cells Chondrocyte | [83] [84] [71] [85] |
| CD63 | Hepatocellular carcinoma (HepG2) CD8+ T-cells | [86] [87] |
| Chemical engineering | ||
| Covalent reactions | Glioma Cerebral vascular endothelial cell | [88] [12] |
| Noncovalent reactions | Hepatoma 22 subcutaneous cancer cells Cancer cells (SKOV3, HCC-1954) | [89,90] [20] |
Physical engineering
Physical methods mainly include membrane fusion [Figure 3A] and membrane coating [Figure 3B]. The liposome mediated MEVs and BEVs membrane fusion strategy is an important engineering approach that endows EVs with specific functional ligands. Liposomes with targeting molecules on the surface can be delivered into EVs through membrane fusion. A mixture of MEVs and liposomes incubated at 37 °C for 12 h could form hybrid nanocarriers. Lin et al.[79] developed MEV-liposome hybrid nanoparticles to accurately deliver large plasmids, such as CRISPR-Cas9, into mesenchymal stem cells (MSCs). Similarly, our team constructed MEV-liposome hybrid nanoparticles with the ability to target bone through spontaneous membrane fusion[18]. Yang et al.[91] reported that virus-mimetic fusogenic MEVs could deliver membrane proteins to the target cell membrane by membrane fusion. Gao et al.[92] also developed a virus-mimicking fusogenic vesicle with fusogenic proteins that could target sialic-acid-containing receptors on MEVs and promote membrane fusion. In addition, the fusion of MEVs derived from different cells and functionalized liposomes could be triggered by polyethylene glycol (PEG)[93].
On the other hand, membrane coating is a promising nanotechnology for disease-relevant targeting. The biological characteristics of cell membranes endow nanoparticles with broader applications, including targeting ability[94]. Various cell types, including mammalian cells (such as red blood cells[95], platelet[96], and cancer cells[97]) and bacterial cells[40,98], have been used for membrane sources. The bacterial membrane could be used for vaccination because of the immunogenic caused by peptidoglycan and outer membrane proteins[99]. Recently, BEV-coated multi-antigenic nano-vaccines have been developed. BEV coating and indocyanine green (ICG)-loaded magnetic mesoporous silica nanoparticles (MSN) were developed by Chen et al.[80] to regulate antigen presentation pathways in dendritic cells. The in vivo data show that the BEV-ICG-MSNs vaccine could target lymph nodes from the injection site[80].
Biological engineering
We can use biological engineering to fuse the gene sequence of the protein with the gene sequence of the selected membrane protein[75]. The most commonly used methods of biological engineering are LAMP-2B [Figure 3C][71,83,85,100,101] and CD63 [Figure 3D][86,87,102]. LAMP-2B, a member of the lysosome-associated membrane protein family, is the most widely used MEV membrane protein for displaying targeting motifs. The N-terminus of LAMP-2B is present on the outer surface of MEVs, and any targeting sequences can be added [Figure 3B]. Alvarez-Erviti et al.[83] used EVs derived from dendritic cells containing Lamp2B-RVG (neuron-specific rabies viral glycoprotein) to achieve neuronal cell (Neuro-2a) targeting. To obtain colorectal tumor (HCT-116) targeting abilities, Liang et al.[84] fused a human epidermal growth factor receptor 2 (HER2) affibody to the N-terminus of LAMP-2B. Xu et al.[71] fused peptide E7 and LAMP-2B to produce MEVs with the ability to target synovial fluid-derived MSCs. By fusing a chondrocyte-affinity peptide with LAMP-2B, Liang et al.[85] generated chondrocyte-targeting MEVs. The transmembrane protein CD63 can also be used to display targeting sequences [Figure 3C]. Engineered MEVs with hepatocellular carcinoma (HepG2)-targeting ability were developed by expression in 293T cell hosts and gene fusion between the CD63 and ApoA-1 sequences[86]. For targeting CD8+ T cells, Kanuma et al.[87] constructed engineered MEVs by fusing ovalbumin (OVA) antigen to CD63.
Chemical engineering
Chemical engineering is another common method that allows various ligands to be displayed in the membrane by covalent reactions [Figure 3E][103-105] or noncovalent reactions [Figure 3F][106-108]. The most applied covalent reactions include click chemistry bioconjugation and aldehyde amine condensation. Click chemistry has been the most used method for attaching targeting peptides to the surface of MEVs in recent years[12,88,109]. Sulfhydryl groups, widely present in membrane proteins, can react with maleimide by Michael addition reaction, which is usually used to selectively modify protein sites. Therefore, various functional molecules are added to the surface of MEVs by conjugating sulfhydryl groups. Jia et al.[88] and Tian et al.[12] also applied click chemistry to develop targeting MEVs, which have glioma-targeting and cerebral vascular endothelial cell-targeting abilities, respectively. In addition, the binding of the anchor peptides CP05 and CD63 via covalent bonding is an example of bioconjugation, indicating that engineered MEVs have emerging prospects in targeted therapy[103]. Tran et al.[16] reported that the combination of aptamers and molecularly targeted MEVs is an intelligent engineering nanovesicle for precision medicine. Moreover, MEVs could be conjugated to aptamers by N-ethyl-N’-[3-(dimethylamino) propyl] carbodiimide/N-hydroxysuccinimide amidation and aldehyde amine condensation reactions[82].
The most commonly applied noncovalent reactions include hydrophobic insertion, multivalent electrostatic interactions, and receptor-ligand binding [Figure 3E]. Lipids or amphiphilic molecules can be inserted into the lipid bilayer of MEVs, and the hydrophilic part is displayed on the exterior. The hydrophobic insertion can be easily accomplished with cells and MEVs at different temperatures. The commercial amphiphilic molecule DSPE-PEG can couple with ligands such as aminoethylanisamide (AA)[110], RGD[107,111], folate[112], etc., to enhance the targeting ability of MEVs. Multivalent electrostatic interactions and receptor-ligand binding are less frequently applied to MEVs. Nakase et al.[113] utilized the negatively charged characteristics of MEVs to bind cationic lipids, which promoted the formation of pH-sensitive fusion peptides and MEVs. Qi et al.[89,90] constructed targeted and magnetic MEVs by receptor-ligand binding. These researchers coincubated reticulocyte-derived EVs (containing transferrin receptors on the membrane surface) and transferrin-conjugated multiple superparamagnetic iron oxide nanoparticles. Importantly, Gujrati et al.[20] used this method to construct engineered BEVs with an anti-HER2 affibody on the outer membrane surface. The engineered BEVs could target and kill cancer cells without nonspecific side effects. Although research on BEVs is not as developed as that on MEVs, their membrane structures are similar. Therefore, the engineering techniques of MEVs provide a good foundation for the in-depth study of BEVs in the future.
THE APPLICATION OF ENGINEERED MEVS AND BEVS IN TARGETED THERAPY
We summarize above the biogenesis, isolation, and characterization methods of MEVs and BEVs, as well as the various engineering methods, and engineered MEVs and BEVs can be used for targeted therapy in a variety of tissues. Next, we summarize the applications of engineered MEVs and BEVs with an enhanced targeting ability in tumors, brain, and bone diseases [Figure 4 and Table 2].
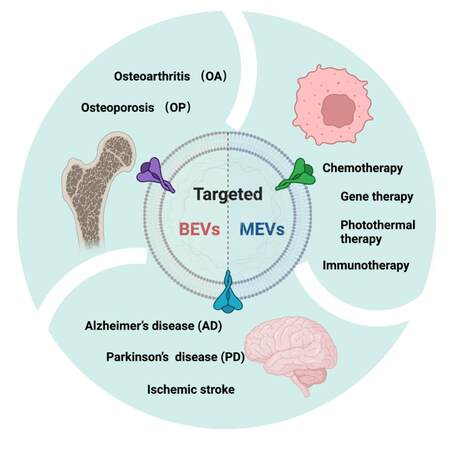
Figure 4. The application of engineered MEVs and BEVs in targeted therapy. The figure was created with Biorender.com. MEVs: Mammalian extracellular vesicles; BEVs: bacterial extracellular vesicles.
Summary of the application of engineered MEVs and BEVs in cancer, brain, and bone disease
| Disease/Therapy | EV source | Ref. |
| Cancer | ||
| Chemotherapy | Mouse immature DCs Serum MDA231/B16F10 cells | [100] [89] [114] |
| Gene therapy | HEK293T Mouse 4 T1 cells E. coli K-12 W3110 | [101] [115] [20] |
| Photothermal therapy | 4T1/SKBR3/HepG2 cells MCF-7 cells E. coli K-12 W3110 | [116] [111] [15,117] |
| Immunotherapy | CAR-T cells HEK-293T cells | [118] [119] |
| Brain | ||
| Alzheimer’s disease Parkinson’s disease Ischemic stroke | Dendritic cells Mesenchymal stem cells Dendritic cells HEK293T cells HEK293T cells HEK293T Mesenchymal stem cells | [83] [120] [121] [81] [122] [123] [124] |
| Bone | ||
| Osteoarthritis Osteoporosis | Mesenchymal stem cells Chondrocyte Endothelial cells NIH-3T3 cells. | [71] [85] [17] [18] |
Cancer
Various natural nanoscale MEVs and BEVs have been applied as drug delivery nanocarriers in cancer therapy[125-129]. The engineered targeting MEVs and BEVs show enhanced therapeutic effects for future cancer therapy, including chemotherapy, gene therapy, photothermal therapy, and immunotherapy.
Chemotherapy
The treatment of tumor diseases routinely involves chemotherapeutic drugs, but chemotherapy drugs do not show specific targeting and have significant cytotoxic side effects, resulting in poor therapeutic effects. Targeted drug delivery based on engineered MEVs and BEVs could increase the local concentration and minimize cytotoxic side effects, consequently improving efficacy. Tian et al.[100] constructed iRGD-MEVs for the delivery of doxorubicin to the breast cancer cell line MDA-MB-231. The chemotherapeutic drug doxorubicin was encapsulated in targeting iRGD-MEVs by electroporation technology. Intravenous injection of iRGD-MEVs specifically delivered doxorubicin to tumor tissues and resulted in inhibition of tumor growth. Qi et al.[89] developed dual-functional (magnetic and targeting ability) MEVs loaded with doxorubicin to target hepatoma 22 subcutaneous cancer cells. Dual-functional MEVs enhanced the cancer-targeting ability under a magnetic field and suppressed tumor growth [Figure 5A]. Similarly, A33 antibody functionalized MEVs with doxorubicin were used to target the colorectal cancer cell line LIM1215[114].
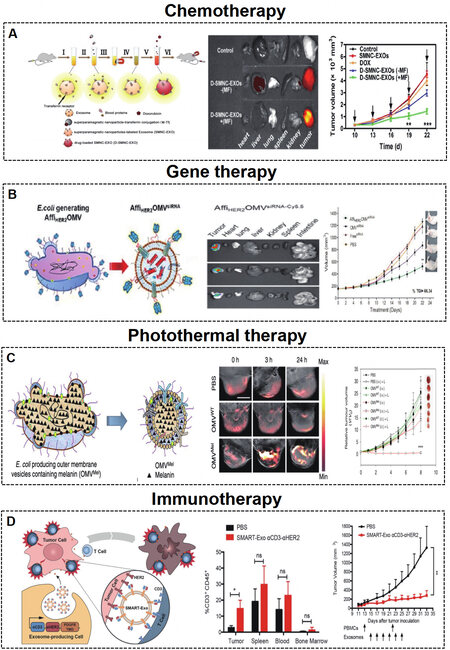
Figure 5. (A) Engineered MEVs for tumor chemotherapy. Schematic illustration of the construction and delivery of doxorubicin loaded in MEVs, which show tumor targeting and antitumor effects[89]. Copyright 2016 American Chemical Society. (B) Engineered BEVs for tumor gene therapy. Schematic illustration of the construction and delivery of siRNA loaded in BEVs, which show tumor targeting and antitumor effects[20]. Copyright 2014 American Chemical Society. (C) Engineered MEVs for tumor photothermal therapy[15]. Schematic illustration of the construction of BEV-Mel, which shows tumor targeting and antitumor effects. Copyright 2019 Springer Nature. (D) Engineered MEVs for tumor immunotherapy[119]. Schematic illustration of the construction of SMART-MEVs, which show tumor targeting and antitumor effects. Copyright 2020 Elsevier. MEVs: Mammalian extracellular vesicles; BEVs: bacterial extracellular vesicles; SMART-MEVs: synthetic multivalent antibodies retargeted MEVs. Significance of finding was defined as follows: not significant, nsP > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001.
Gene therapy
Gene therapy is a strategy to correct or compensate for abnormal gene expression in tumor cells by delivering nucleic acids such as siRNAs, miRNAs, etc., to achieve the purpose of treatment, which has proven to be a promising cancer treatment approach[130]. Specifically, EVs can protect RNA from degradation, which ensures the stability and bioactivity of RNA after targeting cells[131]. Bai et al.[101] reported engineered targeting tLyp-1 MEVs for efficient delivery of SOX2 siRNA to HEK293T cells. The engineered tLyp-1 MEVs had high transfection efficiency in non-small-cell lung cancer (NSCLC) and a high SOX2 gene silencing ability in NSCLC stem cells. Zhao et al.[115] exploited biomimetic CBSA-MEV nanoparticles loaded with S100A4 siRNA, which effectively targeted the lung and showed excellent gene-silencing effects. Moreover, bioengineered BEVs have been used for targeted therapy of tumors. Gujrati et al.[20] constructed BEVs with low immunogenicity that can target cancer cells by delivering KSP siRNA [Figure 5B].
Photothermal therapy
The problems of recurrence, drug toxicity, and multidrug resistance are still difficult to overcome with traditional surgical intervention, chemotherapy, and gene therapy. Photothermal therapy is a nontoxic and noninvasive tumor-targeted treatment method[111,116,132,133]. The combination of engineered bioactive material loaded EVs and photothermal therapy is a promising method for cancer therapy. Bose et al.[116] developed MEVs loaded with anti-miRNA-21-coated gold-iron oxide nanoparticles (GIONs). MEV-GIONs showed strong T2 contrast in magnetic resonance imaging and photothermal effects in breast cancer 4T1 cells. Cao et al.[111] constructed Arg-Gly-Asp (RGD) peptide-MEVs coated with vanadium carbide quantum dots. The resulting MEVs could target cancer cells and access the nucleus to induce low-temperature photothermal therapy, which showed effective tumor destruction. In addition, Mel is highly suitable for photothermal therapy due to its good photothermal conversion efficiency[134,135]. Gujrati et al.[15,117] introduced engineered BEVs derived from E. coli W3110△msbB to carry Mel [Figure 6C]. The engineered BEV-Mel is an excellent anticancer therapy due to its targeting ability, biocompatibility, and scalability. Importantly, BEV-Mel did not induce chronic systemic toxicity or side effects [Figure 5C].
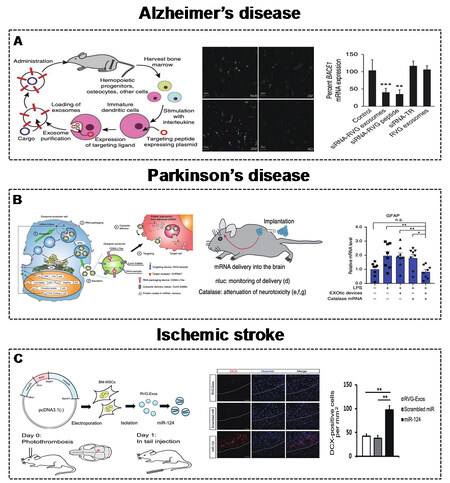
Figure 6. (A) Engineered MEVs for AD. Schematic illustration of the construction of MEVs to deliver BACE1 mRNA with targeting and anti-AD effects[83]. Copyright 2011 Springer Nature. (B) Engineered MEVs for PD. Schematic illustration of the construction of MEVs to deliver catalase mRNA with targeting and anti-PD effects[81]. Copyright 2018 Springer Nature. (C) Engineered MEVs for ischemic stroke.Schematic illustration of the construction of MEVs to deliver catalase miR-124 with targeting and anti-ischemic stroke effects[124]. Copyright 2017 Elsevier. MEVs: Mammalian extracellular vesicles. AD: Alzheimer’s disease. PD: Parkinson’s disease. Significance of finding was defined as follows: not significant, nsP > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001.
Immunotherapy
Immunotherapy is another promising method in the field of tumor therapy[119,136]. Precision targeted therapy with chimeric antigen receptor T (CAR-T) cells is a new type of tumor treatment that has achieved good results in clinical tumor treatment. Fu et al.[118] introduced engineered MEVs derived from CAR-T cells. CAR-containing MEVs could express many cytotoxic molecules and target and kill cancer cells. Shi et al.[119] developed synthetic multivalent antibodies retargeted MEVs (SMART-MEVs), which could specifically target CD3-positive T cells and HER-2 breast cancer cells. The SMART-MEVs exhibited valid and specific antitumor effects [Figure 5D].
Brain
In addition to their strong loading capacity, low toxicity, and low immunogenicity, EVs can also cross the blood-brain barrier. Therefore, EVs can be widely used as a therapeutic vehicle for brain and neurodegenerative diseases[13,137,138]. Here, we summarize the therapeutic effects of engineered targeting EVs in brain diseases, such as AD, PD, and ischemic stroke.
Alzheimer’s disease
AD is a progressive neurodegenerative disease involving the superfluous accumulation of β-amyloid, which is produced by the BACE1 protein[139]. Therefore, controlling the expression of the BACE1 protein is an effective way to control AD. Alvarez-Erviti et al.[83] described engineered RVG (central nervous system-specific peptide)-MEVs (derived from DCs) to specifically deliver GAPDH siRNA to neurons, oligodendrocytes, and microglia in the brain. Mice were injected intravenously with RVG-MEVs, and the engineered MEVs resulted in a significant decrease in the expression of BACE1 mRNA and protein [Figure 6A]. Cui et al.[120] also demonstrated that intravenously infused RVG-MEVs (derived from MSCs) show strong targeting to the cortex and hippocampus, effectively improving learning and memory abilities.
Parkinson’s disease
PD is another progressive neurodegenerative disease that involves the formation of Lewy bodies, which is affected by excessive accumulation of α-synuclein (α-Syn)[140-142]. Similarly, decreasing α-synuclein in brain cells could delay PD. Cooper et al.[121] delivered α-Syn siRNA by RVG-MEVs (derived from murine dendritic cells) to reduce α-Syn accumulation in the brain. Kojima et al.[81] developed MEVs with targeting, cytoplasmic delivery capabilities, and specific RNA encapsulation by EV production booster devices. The delivery of therapeutic catalase mRNA significantly alleviated neurotoxicity and neuroinflammation in mice [Figure 6B]. Liu et al.[122] also modified the membrane surface with the RVG peptide for the targeting ability of MEVs, which delivered MOR siRNA to Neuro2A cells in the brain, leading to decreased morphine addiction.
Ischemic stroke
Ischemic stroke is a disease caused by cerebral arterial stenosis that releases high-mobility group box 1 (HMGB1) to the extracellular spaces and results in inflammatory reactions[143]. The knockdown of HMGB1 in the brain may be an effective anti-inflammatory strategy to improve ischemic stroke. Kim[123] applied brain-targeting RVG-MEVs (derived from HEK293T cells) to precisely deliver HMGB1 siRNA. HMGB1 siRNA was loaded by electroporation technology. Engineered RVG-MEVs with HMGB1 siRNA successfully reduced the expression of HMGB1 protein and apoptosis levels in the brain. In addition, the delivery of miRNAs such as miR-124 is involved in the neuro-remodeling process[144,145]. Using this strategy, Yang et al.[124] constructed RVG-MEVs to deliver miR-124 to the infarct site and protect against ischemic injury [Figure 6C].
Bone
Bone is an internal support system that provides the structural foundation for the human body and muscle[146-149]. The most common bone diseases, such as OA, OP, bone fractures, and bone defects, have been linked to MEVs and BEVs[150-153]. The application of engineering techniques to enhance the bone targeting ability of EVs has substantially increased their therapeutic efficacy in these bone-related diseases. Conventional fractures or bone defects often require biomaterials, such as hydrogels[154-156] and scaffolds[157-160], for therapeutic effects. Here, we review the application of targeted EVs in OA and OP.
Osteoarthritis
OA is a common joint disease with no recognized mechanism[161]. Cartilage degeneration, subchondral bone sclerosis, and synovial inflammation are prominent features of OA[162,163]. There are no effective OA treatments approved by official agencies, except for joint replacement. Although the precise mechanism of OA is still unclear, EVs, especially targeted EVs, play a vital role during the progression of OA, indicating their exciting therapeutic prospects[164]. The small molecule drug kartogenin (KGN) was shown to induce synovial fluid-derived MSCs (SF-MSCs) to differentiate into chondrocytes[165-167]. Xu et al.[71] reported the targeted delivery of KGN to SF-MSCs by engineered MEVs to accelerate chondrogenesis [Figure 7A]. The targeting ability of engineered MEVs is due to E7-Lamp2B. KGN was loaded inside MEVs by electroporation. Moreover, miR-140 is regarded as a promising agent for the treatment of OA due to its dual roles in both homeostasis and cartilage[168,169]. Liang et al.[85] reported a similar targeted strategy to deliver miR-140 to chondrocytes by engineered MEVs to alleviate the progression of OA. The targeting ability of engineered MEVs is due to CAP-Lamp2B. miR-140 was also introduced into MEVs by electroporation.
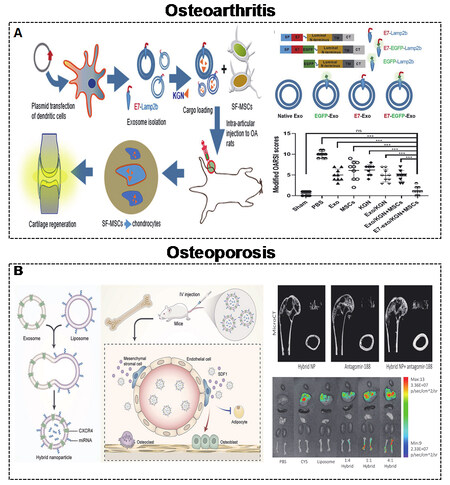
Figure 7. (A) Engineered MEVs for OA. Schematic illustration of the construction of MEVs to deliver KGN with targeting and anti-OA effects[71]. Copyright 2021 Springer Nature. (B) Engineered MEVs for OP. Schematic illustration of the construction of hybrid nanoparticles to deliver antagomir-188 with targeting and anti-OP effects[18]. Copyright 2021 Elsevier. MEVs: Mammalian extracellular vesicles; OA: osteoarthritis; KGN: kartogenin; OP: osteoporosis. Significance of finding was defined as follows: not significant, nsP > 0.05; ***P < 0.0001.
Osteoporosis
OP is a systemic metabolic disease of the skeletal system characterized by fragility fracture[170-172]. The main cause of OP is an imbalance in the metabolism of osteoblasts and osteoclasts. Postmenopausal women suffer from osteoporosis-related fractures throughout their lifetime[173]. Bone-targeted EVs are optimal interventions to improve postmenopausal OP (PMO). Our group demonstrated that MEVs derived from endothelial cells have a better targeting ability than those from osteoblasts[17]. In addition, our team developed targeting MEVs by displaying C-X-C motif chemokine receptor 4 (CXCR4) on their surface[18]. miR-188 was shown to inhibit osteogenesis of bone marrow mesenchymal stem cells (BMSCs), and the knockdown of miR-188 also improved bone loss[174]. Therefore, we combined CXCR4+ with liposomes containing antagomir-188 to generate hybrid nanoparticles for OP treatment [Figure 7B].
CONCLUSION AND PERSPECTIVE
Over time, the pivotal role of EVs in cell-to-cell communications, in contrast to their initial roles as “garbage bags”, has been established. Nanosized EVs have many advantages, such as rich functional contents, a stable membrane structure, good biocompatibility, low immunogenicity, etc. Such cell membrane-derived vesicles have been explored in prokaryotic and eukaryotic cells[95,175,176]. As the two most common types of EVs in the biomedical field, MEVs and BEVs have been studied, resulting in major progress in the biogenesis, isolation, and characterization of these vesicles. For biogenesis, MEVs are formed by endosomal systems including sequential plasma membrane invagination and membrane fusion of eukaryotic cells. BEVs are generated by endolysin-triggered cell lysis and membrane blebbing of bacteria. The isolation and characterization of MEVs and BEVs determine their further application in biomedicine. Here, an ultracentrifugation-based isolation protocol is described for MEVs. An effective isolation method based on ultracentrifugation and density gradient centrifugation is also described for BEVs. For characterization, TEM and NTA are commonly used to characterize the sizes, shapes, and concentrations of these vesicles. However, different types of MEVs contain the same protein molecules, such as TSG101, CD63, and CD81, which are always used as specific markers by WB. In contrast, although several membrane proteins, such as OmpF and OmpA, have been used for the characterization of E. coli EVs, specific markers of BEVs are still a major challenge.
The use of MEVs and BEVs also has many challenges, such as poor targeting specificity. Targeted drug delivery of EVs was proposed in 2011 and has since received increasing attention due to their excellent characteristics[177]. Targeted modification methods have been applied in MEVs and BEVs to increase the targeting ability and healing efficacy. Targeted engineering aims to increase the local concentration of EVs at diseased sites, thereby reducing toxicity and side effects and maximizing healing efficacy. Both MEVs and BEVs are lipid bilayer-enclosed structures containing various biomolecules. Therefore, most engineering methods are universal. Here, physical, biological, and chemical engineering methods based on membranes to modify MEVs are described, which could also guide the modification of BEVs. Furthermore, the applications of engineered MEVs and BEVs in targeted therapy, such as therapy for tumors and brain and bone diseases, are summarized. MEVs and BEVs with targeting capabilities are usually administered systemically intravenously, and they will accumulate at the target site for better therapeutic efficiency. In addition to the direct injection, the incorporation of MEVs and BEVs with biomaterials is also a promising approach to enhance the healing efficacy[160]. Different types of biomaterials, such as hydrogels[154,178-181] and scaffolds[157,159,182,183], have been developed to achieve high retention rates of EVs and healing efficacy of tissue. In situ injection is another targeted technique, allowing MEVs and BEVs to act directly at the injured tissues. However, some hard-to-reach tissues still require targeted EVs. The many big deals recently by large pharmaceutical companies indicate that the industry expects MEVs and BEVs to deliver drugs to hard-to-reach tissues[184].
Although MEVs have been more extensively studied than BEVs, one of the challenges of MEVs is the limited yield. BEVs are easily available due to the rapid proliferative abilities, mature culture methods, and gene editing techniques of bacteria[185,186]. In addition, the scalability, low cost, and environmental friendliness of bacterial fermentation culture indicate that the industrialization of BEVs is possible[187,188]. Synthetic biology can also be used to confer additional functions on bacteria and their associated BEVs[189,190]. Moreover, several biotherapeutic bacteria, especially human commensal bacteria, such as E. coli Nissle 1917[48], A. muciniphila[191], and L. rhamnosus GG[192], are being investigated in clinical trials[193,194]. Therefore, BEVs derived from probiotics are promising pharmaceutical agents in the biomedical field. Importantly, BEVs are safe because they are cell-free. Both oral and intravenous BEVs were well tolerated and resulted in low immunogenic responses[15,19,20]. Therefore, the topic of “nonmammalian EVs, especially BEVs” is receiving more attention in the latest MISEV. The mature application system of MEVs can also lay a solid foundation for BEVs in biomedical fields. Studies on MEVs and BEVs can inspire each other and draw important elements from each type to enhance functional and therapeutic effcacy. Overall, the rise of targeted therapeutics of engineered MEVs and BEVs shows promise for future clinical translation of EVs.
DECLARATIONS
Authors’ contributions
Liu H and Geng Z contributed equally to this work
Drafted the manuscript: Liu H, Geng Z
Reviewed the manuscript: Liu H, Geng Z, Su J
Approved the submitted manuscript: Su J
All authors have read and agreed to the published version of the manuscript.
Availability of data and materials
Not applicable.
Financial support and sponsorship
This work was supported by the National Key R&D Program of China (2018YFC2001500); National Natural Science Foundation of China (NSFC) Key Research Program in Aging (91749204); National Natural Science Foundation of China (82172098, 81771491, 81972254).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2022.
REFERENCES
1. Witwer KW, Goberdhan DC, O’Driscoll L, et al. Updating MISEV: evolving the minimal requirements for studies of extracellular vesicles. J Extracell Vesicles 2021;10:e12182.
3. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement Of The International Society For Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles 2018;7:1535750.
4. Liu H, Zhang Q, Wang S, et al. Bacterial extracellular vesicles as bioactive nanocarriers for drug delivery: advances and perspectives. Bioact Mater 2022;14:169-81.
5. O’Brien K, Breyne K, Ughetto S, Laurent LC, Breakefield XO. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat Rev Mol Cell Biol 2020;21:585-606.
6. Herrmann IK, Wood MJA, Fuhrmann G. Extracellular vesicles as a next-generation drug delivery platform. Nat Nanotechnol 2021;16:748-59.
7. Liu C, He D, Cen H, et al. Nucleic acid functionalized extracellular vesicles as promising therapeutic systems for nanomedicine. EVCNA 2022; doi: 10.20517/evcna.2021.21.
8. Duan L, Xu L, Xu X, et al. Exosome-mediated delivery of gene vectors for gene therapy. Nanoscale 2021;13:1387-97.
9. Li M, Zhou H, Yang C, et al. Bacterial outer membrane vesicles as a platform for biomedical applications: an update. J Control Release 2020;323:253-68.
10. Sharma S, Masud MK, Kaneti YV, et al. Extracellular vesicle nanoarchitectonics for novel drug delivery applications. Small 2021;17:e2102220.
11. Jiang C, Fu Y, Liu G, et al. Multiplexed profiling of extracellular vesicles for biomarker development. Nanomicro Lett 2021;14:3.
12. Tian T, Zhang HX, He CP, et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy. Biomaterials 2018;150:137-49.
13. Xu M, Feng T, Liu B, et al. Engineered exosomes: desirable target-tracking characteristics for cerebrovascular and neurodegenerative disease therapies. Theranostics 2021;11:8926-44.
14. Liao Z, Liu H, Ma L, et al. Engineering extracellular vesicles restore the impaired cellular uptake and attenuate intervertebral disc degeneration. ACS Nano 2021;15:14709-24.
15. Gujrati V, Prakash J, Malekzadeh-Najafabadi J, et al. Bioengineered bacterial vesicles as biological nano-heaters for optoacoustic imaging. Nat Commun 2019;10:1114.
16. Tran PHL, Xiang D, Tran TTD, et al. Exosomes and nanoengineering: a match made for precision therapeutics. Adv Mater 2020;32:e1904040.
17. Song H, Li X, Zhao Z, et al. Reversal of osteoporotic activity by endothelial cell-secreted bone targeting and biocompatible exosomes. Nano Lett 2019;19:3040-8.
18. Hu Y, Li X, Zhang Q, et al. Exosome-guided bone targeted delivery of Antagomir-188 as an anabolic therapy for bone loss. Bioact Mater 2021;6:2905-13.
19. Kim OY, Park HT, Dinh NTH, et al. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat Commun 2017;8:626.
20. Gujrati V, Kim S, Kim SH, et al. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano 2014;8:1525-37.
21. Kalluri R, LeBleu VS. function, and biomedical applications of exosomes. Science 2020;367:eaau6977.
22. Kim J, Song Y, Park CH, Choi C. Platform technologies and human cell lines for the production of therapeutic exosomes. EVCNA 2021; doi: 10.20517/evcna.2020.01.
23. Work E, Knox KW, Vesk M. The chemistry and electron microscopy of an extracellular lipopolysaccharide from escherichia coli. Ann N Y Acad Sci 1966;133:438-49.
24. Knox KW, Vesk M, Work E. Relation between excreted lipopolysaccharide complexes and surface structures of a lysine-limited culture of Escherichia coli. J Bacteriol 1966;92:1206-17.
25. Schwechheimer C, Kuehn MJ. Outer-membrane vesicles from Gram-negative bacteria: biogenesis and functions. Nat Rev Microbiol 2015;13:605-19.
26. Brown L, Wolf JM, Prados-Rosales R, Casadevall A. Through the wall: extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat Rev Microbiol 2015;13:620-30.
27. Harding C, Heuser J, Stahl P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J Cell Biol 1983;97:329-39.
28. Pan B, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes. in vitro ;33:967-78.
30. Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med 1996;183:1161-72.
31. Prangishvili D, Holz I, Stieger E, et al. Sulfolobicins, specific proteinaceous toxins produced by strains of the extremely thermophilic archaeal genus Sulfolobus. J Bacteriol 2000;182:2985-8.
32. Valadi H, Ekström K, Bossios A, et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007;9:654-9.
33. Fonseka P, Marzan AL, Mathivanan S. Introduction to the Community of Extracellular Vesicles. Subcell Biochem 2021;97:3-18.
34. Mathieu M, Martin-Jaular L, Lavieu G, Théry C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol 2019;21:9-17.
35. Kowal J, Arras G, Colombo M, et al. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc Natl Acad Sci U S A 2016;113:E968-77.
36. Kowal J, Tkach M, Théry C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol 2014;29:116-25.
37. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol 2013;200:373-83.
38. Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008;319:1244-7.
39. Palomino RA, Vanpouille C, Costantini PE, Margolis L. Microbiota-host communications: Bacterial extracellular vesicles as a common language. PLoS Pathog 2021;17:e1009508.
40. Wang S, Gao J, Wang Z. Outer membrane vesicles for vaccination and targeted drug delivery. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2019;11:e1523.
41. Garcia-Martin R, Wang G, Brandão BB, et al. MicroRNA sequence codes for small extracellular vesicle release and cellular retention. Nature 2022;601:446-51.
42. Díaz-Garrido N, Badia J, Baldomà L. Microbiota-derived extracellular vesicles in interkingdom communication in the gut. J Extracell Vesicles 2021;10:e12161.
43. Bittel M, Reichert P, Sarfati I, et al. Visualizing transfer of microbial biomolecules by outer membrane vesicles in microbe-host-communication. in vivo ;10:e12159.
44. Vanaja SK, Russo AJ, Behl B, et al. Bacterial outer membrane vesicles mediate cytosolic localization of LPS and caspase-11 activation. Cell 2016;165:1106-19.
45. Deo P, Chow SH, Han ML, et al. Mitochondrial dysfunction caused by outer membrane vesicles from gram-negative bacteria activates intrinsic apoptosis and inflammation. Nat Microbiol 2020;5:1418-27.
46. Somerville JE Jr, Cassiano L, Darveau RP. Escherichia coli msbB gene as a virulence factor and a therapeutic target. Infect Immun 1999;67:6583-90.
47. Grozdanov L, Raasch C, Schulze J, et al. Analysis of the genome structure of the nonpathogenic probiotic escherichia coli strain nissle 1917. J Bacteriol 2004;186:5432-41.
48. Yan X, Liu XY, Zhang D, et al. Construction of a sustainable 3-hydroxybutyrate-producing probiotic escherichia coli for treatment of colitis. Cell Mol Immunol 2021;18:2344-57.
49. Praveschotinunt P, Duraj-Thatte AM, Gelfat I, et al. Engineered E. coli Nissle 1917 for the delivery of matrix-tethered therapeutic domains to the gut. Nat Commun 2019;10:5580.
50. Domingues S, Nielsen KM. Membrane vesicles and horizontal gene transfer in prokaryotes. Curr Opin Microbiol 2017;38:16-21.
51. Gilmore WJ, Johnston EL, Zavan L, Bitto NJ, Kaparakis-Liaskos M. Immunomodulatory roles and novel applications of bacterial membrane vesicles. Mol Immunol 2021;134:72-85.
52. Lee HJ. Microbe-host communication by small RNAs in extracellular vesicles: vehicles for transkingdom RNA transportation. Int J Mol Sci 2019;20:1487.
53. Dauros-Singorenko P, Blenkiron C, Phillips A, Swift S. The functional RNA cargo of bacterial membrane vesicles. FEMS Microbiol Lett 2018:365.
54. Kumar SR, Kimchi ET, Manjunath Y, et al. RNA cargos in extracellular vesicles derived from blood serum in pancreas associated conditions. Sci Rep 2020;10:2800.
55. la Torre Gomez C, Goreham RV, Bech Serra JJ, Nann T, Kussmann M. “Exosomics”-a review of biophysics, biology and biochemistry of exosomes with a focus on human breast milk. Front Genet 2018;9:92.
56. Zhu Q, Cheng L, Deng C, et al. The genetic source tracking of human urinary exosomes. Proc Natl Acad Sci U S A 2021;118:e2108876118.
57. Liu H, Yuan W, Pang Q, Xue C, Yan X. Single-particle analysis of tear fluid reveals abundant presence of tissue factor-exposing extracellular vesicles with strong coagulation activity. Talanta 2022;239:123089.
58. Lin Y, Dong H, Deng W, et al. Evaluation of salivary exosomal chimeric. GOLM1-NAA35 ;25:3035-45.
59. Höög JL, Lötvall J. Diversity of extracellular vesicles in human ejaculates revealed by cryo-electron microscopy. J Extracell Vesicles 2015;4:28680.
60. Han X, Lee A, Huang S, Gao J, Spence JR, Owyang C. Lactobacillus rhamnosus GG prevents epithelial barrier dysfunction induced by interferon-gamma and fecal supernatants from irritable bowel syndrome patients in human intestinal enteroids and colonoids. Gut Microbes 2019;10:59-76.
61. Liu JH, Chen CY, Liu ZZ, et al. Extracellular vesicles from child gut microbiota enter into bone to preserve bone mass and strength. Adv Sci (Weinh) 2021;8:2004831.
62. Maeki M, Kimura N, Sato Y, Harashima H, Tokeshi M. Advances in microfluidics for lipid nanoparticles and extracellular vesicles and applications in drug delivery systems. Adv Drug Deliv Rev 2018;128:84-100.
63. Zhu L, Xu N, Zhang ZL, Zhang TC. Cell derived extracellular vesicles: from isolation to functionalization and biomedical applications. Biomater Sci 2019;7:3552-65.
64. Hartjes TA, Mytnyk S, Jenster GW, van Steijn V, van Royen ME. Extracellular vesicle quantification and characterization: common methods and emerging approaches. Bioengineering (Basel) 2019;6:7.
65. Li X, Corbett AL, Taatizadeh E, et al. Challenges and opportunities in exosome research-perspectives from biology, engineering, and cancer therapy. APL Bioeng 2019;3:011503.
66. Liang Y, Duan L, Lu J, Xia J. Engineering exosomes for targeted drug delivery. Theranostics 2021;11:3183-95.
67. Li P, Kaslan M, Lee SH, Yao J, Gao Z. Progress in exosome isolation techniques. Theranostics 2017;7:789-804.
68. Gardiner C, Di Vizio D, Sahoo S, et al. Techniques used for the isolation and characterization of extracellular vesicles: results of a worldwide survey. J Extracell Vesicles 2016;5:32945.
69. Xiong Y, Chen L, Yan C, et al. Circulating exosomal miR-20b-5p inhibition restores Wnt9b signaling and reverses diabetes-associated impaired wound healing. Small 2020;16:e1904044.
70. Jiang S, Tian G, Yang Z, et al. Enhancement of acellular cartilage matrix scaffold by Wharton’s jelly mesenchymal stem cell-derived exosomes to promote osteochondral regeneration. Bioact Mater 2021;6:2711-28.
71. Xu X, Liang Y, Li X, et al. Exosome-mediated delivery of kartogenin for chondrogenesis of synovial fluid-derived mesenchymal stem cells and cartilage regeneration. Biomaterials 2021;269:120539.
72. Prados-Rosales R, Brown L, Casadevall A, Montalvo-Quirós S, Luque-Garcia JL. Isolation and identification of membrane vesicle-associated proteins in Gram-positive bacteria and mycobacteria. MethodsX 2014;1:124-9.
73. Chutkan H, Macdonald I, Manning A, Kuehn MJ. Quantitative and qualitative preparations of bacterial outer membrane vesicles. Methods Mol Biol 2013; 966:259-72.
74. Klimentová J, Stulík J. Methods of isolation and purification of outer membrane vesicles from gram-negative bacteria. Microbiol Res 2015;170:1-9.
76. Wang X, Zhang H, Yang H, et al. Cell-derived exosomes as promising carriers for drug delivery and targeted therapy. Curr Cancer Drug Targets 2018;18:347-54.
78. Mathivanan S, Fahner CJ, Reid GE, Simpson RJ. ExoCarta 2012: database of exosomal proteins, RNA and lipids. Nucleic Acids Res 2012;40:D1241-4.
79. Lin Y, Wu J, Gu W, et al. Exosome-liposome hybrid nanoparticles deliver CRISPR/Cas9 system in MSCs. Adv Sci (Weinh) 2018;5:1700611.
80. Chen G, Bai Y, Li Z, et al. Bacterial extracellular vesicle-coated multi-antigenic nanovaccines protect against drug-resistant. Staphylococcus aureus ;10:7131-49.
81. Kojima R, Bojar D, Rizzi G, et al. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson’s disease treatment. Nat Commun 2018;9:1305.
82. Wu P, Zhang B, Ocansey DKW, Xu W, Qian H. Extracellular vesicles: a bright star of nanomedicine. Biomaterials 2021;269:120467.
83. Alvarez-Erviti L, Seow Y, Yin H, et al. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol 2011;29:341-5.
84. Liang G, Zhu Y, Ali DJ, et al. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J Nanobiotechnology 2020;18:10.
85. Liang Y, Xu X, Li X, et al. Chondrocyte-targeted microRNA delivery by engineered exosomes toward a cell-free osteoarthritis therapy. ACS Appl Mater Interfaces 2020;12:36938-47.
86. Liang G, Kan S, Zhu Y, et al. Engineered exosome-mediated delivery of functionally active miR-26a and its enhanced suppression effect in HepG2 cells. Int J Nanomedicine 2018;13:585-99.
87. Kanuma T, Yamamoto T, Kobiyama K, et al. CD63-mediated antigen delivery into extracellular vesicles via DNA vaccination results in robust CD8+ T cell responses. J Immunol 2017;198:4707-15.
88. Jia G, Han Y, An Y, et al. NRP-1 targeted and cargo-loaded exosomes facilitate simultaneous imaging and therapy of glioma. in vitro ;178:302-16.
89. Qi H, Liu C, Long L, et al. Blood exosomes endowed with magnetic and targeting properties for cancer therapy. ACS Nano 2016;10:3323-33.
90. Yang L, Han D, Zhan Q, et al. Blood TfR+ exosomes separated by a pH-responsive method deliver chemotherapeutics for tumor therapy. Theranostics 2019;9:7680-96.
91. Yang Y, Hong Y, Nam GH, et al. Virus-mimetic fusogenic exosomes for direct delivery of integral membrane proteins to target cell membranes. Adv Mater 2017;29:1605604.
92. Gao X, Li S, Ding F, et al. Rapid detection of exosomal micrornas using virus-mimicking fusogenic vesicles. Angew Chem Int Ed Engl 2019;58:8719-23.
93. Piffoux M, Silva AKA, Wilhelm C, Gazeau F, Tareste D. Modification of extracellular vesicles by fusion with liposomes for the design of personalized biogenic drug delivery systems. ACS Nano 2018;12:6830-42.
94. Fang RH, Kroll AV, Gao W, Zhang L. Cell membrane coating nanotechnology. Adv Mater 2018;30:e1706759.
95. Hu CM, Zhang L, Aryal S, et al. Erythrocyte membrane-camouflaged polymeric nanoparticles as a biomimetic delivery platform. Proc Natl Acad Sci U S A 2011;108:10980-5.
96. Su T, Huang K, Ma H, et al. Platelet-inspired nanocells for targeted heart repair after ischemia/reperfusion injury. Adv Funct Mater 2019;29:1803567.
97. Rao L, Bu LL, Cai B, et al. Cancer cell membrane-coated upconversion nanoprobes for highly specific tumor imaging. Adv Mater 2016;28:3460-6.
98. Wang S, Gao J, Li M, Wang L, Wang Z. A facile approach for development of a vaccine made of bacterial double-layered membrane vesicles (DMVs). Biomaterials 2018;187:28-38.
99. Pore D, Chakrabarti MK. Outer membrane protein A (OmpA) from shigella flexneri 2a: a promising subunit vaccine candidate. Vaccine 2013;31:3644-50.
100. Tian Y, Li S, Song J, et al. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials 2014;35:2383-90.
101. Bai J, Duan J, Liu R, et al. Engineered targeting tLyp-1 exosomes as gene therapy vectors for efficient delivery of siRNA into lung cancer cells. Asian J Pharm Sci 2020;15:461-71.
102. Rountree RB, Mandl SJ, Nachtwey JM, et al. Exosome targeting of tumor antigens expressed by cancer vaccines can improve antigen immunogenicity and therapeutic efficacy. Cancer Res 2011;71:5235-44.
103. Gao X, Ran N, Dong X, et al. Anchor peptide captures, targets, and loads exosomes of diverse origins for diagnostics and therapy. Sci Transl Med 2018;10:eaat0195.
104. Zuo B, Qi H, Lu Z, et al. Alarmin-painted exosomes elicit persistent antitumor immunity in large established tumors in mice. Nat Commun 2020;11:1790.
105. Fan Z, Xiao K, Lin J, Liao Y, Huang X. Functionalized DNA enables programming exosomes/vesicles for tumor imaging and therapy. Small 2019;15:e1903761.
106. Di H, Zeng E, Zhang P, et al. General approach to engineering extracellular vesicles for biomedical analysis. Anal Chem 2019;91:12752-9.
107. Wang J, Li W, Lu Z, et al. The use of RGD-engineered exosomes for enhanced targeting ability and synergistic therapy toward angiogenesis. Nanoscale 2017;9:15598-605.
108. Wang J, Li W, Zhang L, et al. Chemically edited exosomes with dual ligand Purified by microfluidic device for active targeted drug delivery to tumor cells. ACS Appl Mater Interfaces 2017;9:27441-52.
109. Khongkow M, Yata T, Boonrungsiman S, et al. Surface modification of gold nanoparticles with neuron-targeted exosome for enhanced blood-brain barrier penetration. Sci Rep 2019;9:8278.
110. Kim MS, Haney MJ, Zhao Y, et al. Engineering macrophage-derived exosomes for targeted paclitaxel delivery to pulmonary metastases:. in vitro ;14:195-204.
111. Cao Y, Wu T, Zhang K, et al. Engineered exosome-mediated near-infrared-II region V2C quantum dot delivery for nucleus-target low-temperature photothermal therapy. ACS Nano 2019;13:1499-510.
112. Pi F, Binzel DW, Lee TJ, et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression. Nat Nanotechnol 2018;13:82-9.
113. Nakase I, Futaki S. Combined treatment with a pH-sensitive fusogenic peptide and cationic lipids achieves enhanced cytosolic delivery of exosomes. Sci Rep 2015;5:10112.
114. Li Y, Gao Y, Gong C, et al. A33 antibody-functionalized exosomes for targeted delivery of doxorubicin against colorectal cancer. Nanomedicine 2018;14:1973-85.
115. Zhao L, Gu C, Gan Y, et al. Exosome-mediated siRNA delivery to suppress postoperative breast cancer metastasis. J Control Release 2020;318:1-15.
116. Bose RJC, Uday Kumar S, Zeng Y, et al. Tumor cell-derived extracellular vesicle-coated nanocarriers: an efficient theranostic platform for the cancer-specific delivery of Anti-miR-21 and imaging agents. ACS Nano 2018;12:10817-32.
117. Gujrati V, Ntziachristos V. Bioengineered bacterial vesicles for optoacoustics-guided phototherapy. Methods Enzymol 2021;657:349-64.
118. Fu W, Lei C, Liu S, et al. CAR exosomes derived from effector CAR-T cells have potent antitumour effects and low toxicity. Nat Commun 2019;10:4355.
119. Shi X, Cheng Q, Hou T, et al. Genetically engineered cell-derived nanoparticles for targeted breast cancer immunotherapy. Mol Ther 2020;28:536-47.
120. Cui GH, Guo HD, Li H, et al. RVG-modified exosomes derived from mesenchymal stem cells rescue memory deficits by regulating inflammatory responses in a mouse model of Alzheimer’s disease. Immun Ageing 2019;16:10.
121. Cooper JM, Wiklander PB, Nordin JZ, et al. Systemic exosomal siRNA delivery reduced alpha-synuclein aggregates in brains of transgenic mice. Mov Disord 2014;29:1476-85.
122. Liu Y, Li D, Liu Z, et al. Targeted exosome-mediated delivery of opioid receptor Mu siRNA for the treatment of morphine relapse. Sci Rep 2015;5:17543.
123. Kim M, Kim G, Hwang DW, Lee M. Delivery of high mobility group box-1 sirna using brain-targeting exosomes for ischemic stroke therapy. J Biomed Nanotechnol 2019;15:2401-12.
124. Yang J, Zhang X, Chen X, Wang L, Yang G. Exosome mediated delivery of mir-124 promotes neurogenesis after ischemia. Mol Ther Nucleic Acids 2017;7:278-87.
125. Abhange K, Makler A, Wen Y, et al. Small extracellular vesicles in cancer. Bioact Mater 2021;6:3705-43.
126. Zhuang Q, Xu J, Deng D, et al. Bacteria-derived membrane vesicles to advance targeted photothermal tumor ablation. Biomaterials 2021;268:120550.
127. Chronopoulos A, Kalluri R. Emerging role of bacterial extracellular vesicles in cancer. Oncogene 2020;39:6951-60.
129. Han M, Pang B, Zhou C, et al. Liquid biopsy of extracellular vesicle biomarkers for prostate cancer personalized treatment decision. EVCNA 2022; doi: 10.20517/evcna.2021.20.
131. Vader P, Mol EA, Pasterkamp G, Schiffelers RM. Extracellular vesicles for drug delivery. Adv Drug Deliv Rev 2016;106:148-56.
133. Webb JA, Ou YC, Faley S, et al. Theranostic gold nanoantennas for simultaneous multiplexed Raman imaging of immunomarkers and photothermal therapy. ACS Omega 2017;2:3583-94.
134. Stritzker J, Kirscher L, Scadeng M, et al. Vaccinia virus-mediated melanin production allows MR and optoacoustic deep tissue imaging and laser-induced thermotherapy of cancer. Proc Natl Acad Sci U S A 2013;110:3316-20.
135. Fan Q, Cheng K, Hu X, et al. Transferring biomarker into molecular probe: melanin nanoparticle as a naturally active platform for multimodality imaging. J Am Chem Soc 2014;136:15185-94.
136. Cheng Q, Shi X, Han M, et al. Reprogramming exosomes as nanoscale controllers of cellular immunity. J Am Chem Soc 2018;140:16413-7.
137. Xiao L, Hareendran S, Loh YP. Function of exosomes in neurological disorders and brain tumors. Extracell Vesicles Circ Nucl Acids 2021;2:55-79.
138. Cuesta CM, Guerri C, Ureña J, Pascual M. Role of microbiota-derived extracellular vesicles in gut-brain communication. Int J Mol Sci 2021;22:4235.
139. Hampel H, Vassar R, De Strooper B, et al. The β-Secretase BACE1 in Alzheimer’s disease. Biol Psychiatry 2021;89:745-56.
140. Jiang C, Hopfner F, Katsikoudi A, et al. Serum neuronal exosomes predict and differentiate Parkinson’s disease from atypical parkinsonism. J Neurol Neurosurg Psychiatry 2020;91:720-9.
141. Jiang C, Hopfner F, Berg D, et al. Validation of α-Synuclein in L1CAM-immunocaptured exosomes as a biomarker for the stratification of parkinsonian syndromes. Mov Disord 2021;36:2663-9.
142. Fu Y, Jiang C, Tofaris GK, Davis JJ. Facile Impedimetric analysis of neuronal exosome markers in Parkinson’s disease diagnostics. Anal Chem 2020;92:13647-51.
144. Dharap A, Bowen K, Place R, Li LC, Vemuganti R. Transient focal ischemia induces extensive temporal changes in rat cerebral microRNAome. J Cereb Blood Flow Metab 2009;29:675-87.
145. Jeyaseelan K, Lim KY, Armugam A. MicroRNA expression in the blood and brain of rats subjected to transient focal ischemia by middle cerebral artery occlusion. Stroke 2008;39:959-66.
146. Ren X, Liu H, Wu X, et al. Reactive oxygen species (ROS)-responsive biomaterials for the treatment of bone-related diseases. Front Bioeng Biotechnol 2021;9:820468.
147. Chen S, Chen X, Geng Z, Su J. The horizon of bone organoid: a perspective on construction and application. Bioact Mater 2022;18:15-25.
148. Hu Y, Chen X, Wang S, Jing Y, Su J. Subchondral bone microenvironment in osteoarthritis and pain. Bone Res 2021;9:20.
149. Gu J, Zhang Q, Geng M, et al. Construction of nanofibrous scaffolds with interconnected perfusable microchannel networks for engineering of vascularized bone tissue. Bioact Mater 2021;6:3254-68.
150. Puhm F, Boilard E, Machlus KR. Platelet extracellular vesicles: beyond the blood. Arterioscler Thromb Vasc Biol 2021;41:87-96.
151. Wu X, Wang Y, Xiao Y, et al. Extracellular vesicles: potential role in osteoarthritis regenerative medicine. J Orthop Translat 2020;21:73-80.
152. Liu Y, Ma Y, Zhang J, Yuan Y, Wang J. Exosomes: a novel therapeutic agent for cartilage and bone tissue regeneration. Dose Response 2019;17:1559325819892702.
153. Tao SC, Guo SC. Extracellular vesicles in bone: “dogrobbers” in the “eternal battle field”. Cell Commun Signal 2019;17:6.
154. Zhang Y, Xie Y, Hao Z, et al. Umbilical mesenchymal stem cell-derived exosome-encapsulated hydrogels accelerate bone repair by enhancing angiogenesis. ACS Appl Mater Interfaces 2021;13:18472-87.
155. Re F, Gabusi E, Manferdini C, Russo D, Lisignoli G. Bone regeneration improves with mesenchymal stem cell derived extracellular vesicles (EVs) combined with scaffolds: a systematic review. Biology (Basel) 2021;10:579.
156. Pishavar E, Luo H, Naserifar M, et al. Advanced hydrogels as exosome delivery systems for osteogenic differentiation of MSCs: application in bone regeneration. Int J Mol Sci 2021;22:6203.
157. Fan J, Lee CS, Kim S, et al. Generation of small rna-modulated exosome mimetics for bone regeneration. ACS Nano 2020;14:11973-84.
158. Zha Y, Li Y, Lin T, et al. Progenitor cell-derived exosomes endowed with VEGF plasmids enhance osteogenic induction and vascular remodeling in large segmental bone defects. Theranostics 2021;11:397-409.
159. Zhai M, Zhu Y, Yang M, Mao C. Human mesenchymal stem cell derived exosomes enhance cell-free bone regeneration by altering their miRNAs profiles. Adv Sci (Weinh) 2020;7:2001334.
160. Lazar S, Mor S, Chen J, Hao D, Wang A. Bioengineered extracellular vesicle-loaded bioscaffolds for therapeutic applications in regenerative medicine. Extracell Vesicles Circ Nucl Acids 2021;2:175-8.
161. Xue S, Zhou X, Sang W, et al. Cartilage-targeting peptide-modified dual-drug delivery nanoplatform with NIR laser response for osteoarthritis therapy. Bioact Mater 2021;6:2372-89.
163. Martel-Pelletier J, Barr AJ, Cicuttini FM, et al. Osteoarthritis. Nat Rev Dis Primers 2016;2:16072.
164. Ragni E, Orfei CP, Papait A, de Girolamo L. Comparison of miRNA cargo in human adipose-tissue vs. amniotic-membrane derived mesenchymal stromal cells extracellular vesicles for osteoarthritis treatment. EVCNA 2021; doi: 10.20517/evcna.2021.11.
165. Johnson K, Zhu S, Tremblay MS, et al. A stem cell-based approach to cartilage repair. Science 2012;336:717-21.
166. Kang ML, Ko JY, Kim JE, Im GI. Intra-articular delivery of kartogenin-conjugated chitosan nano/microparticles for cartilage regeneration. Biomaterials 2014;35:9984-94.
167. Xu X, Shi D, Shen Y, et al. Full-thickness cartilage defects are repaired via a microfracture technique and intraarticular injection of the small-molecule compound kartogenin. Arthritis Res Ther 2015;17:20.
168. Zhang R, Ma J, Yao J. Molecular mechanisms of the cartilage-specific microRNA-140 in osteoarthritis. Inflamm Res 2013;62:871-7.
169. Miyaki S, Sato T, Inoue A, et al. MicroRNA-140 plays dual roles in both cartilage development and homeostasis. Genes Dev 2010;24:1173-85.
170. Black DM, Rosen CJ. Clinical Practice. Postmenopausal osteoporosis. N Engl J Med 2016;374:254-62.
171. Li X, Wang L, Huang B, et al. Targeting actin-bundling protein L-plastin as an anabolic therapy for bone loss. Sci Adv 2020;6:eabb7135.
172. Zhao H, Li X, Zhang D, et al. Integrative bone metabolomics-lipidomics strategy for pathological mechanism of postmenopausal osteoporosis mouse model. Sci Rep 2018;8:16456.
174. Li CJ, Cheng P, Liang MK, et al. MicroRNA-188 regulates age-related switch between osteoblast and adipocyte differentiation. J Clin Invest 2015;125:1509-22.
175. Gujrati V, Lee M, Ko YJ, et al. Bioengineered yeast-derived vacuoles with enhanced tissue-penetrating ability for targeted cancer therapy. Proc Natl Acad Sci U S A 2016;113:710-5.
176. MacDiarmid JA, Mugridge NB, Weiss JC, et al. Bacterially derived 400 nm particles for encapsulation and cancer cell targeting of chemotherapeutics. Cancer Cell 2007;11:431-45.
177. Gudbergsson JM, Jønsson K, Simonsen JB, Johnsen KB. Systematic review of targeted extracellular vesicles for drug delivery - considerations on methodological and biological heterogeneity. J Control Release 2019;306:108-20.
178. Xue X, Hu Y, Deng Y, Su J. Recent advances in design of functional biocompatible hydrogels for bone tissue engineering. Adv Funct Mater 2021;31:2009432.
179. Chen W, Zhou Z, Chen D, et al. Bone regeneration using MMP-cleavable peptides-based hydrogels. Gels 2021;7:199.
180. Xiong Y, Chen L, Liu P, et al. All-in-one: multifunctional hydrogel accelerates oxidative diabetic wound healing through timed-release of exosome and fibroblast growth factor. Small 2022;18:e2104229.
181. Li L, Yu F, Zheng L, et al. Natural hydrogels for cartilage regeneration: Modification, preparation and application. J Orthop Translat 2019;17:26-41.
182. Zou Y, Huang B, Cao L, Deng Y, Su J. Tailored mesoporous inorganic biomaterials: assembly, functionalization, and drug delivery engineering. Adv Mater 2021;33:e2005215.
183. Geng Z, Sang S, Wang S, et al. Optimizing the strontium content to achieve an ideal osseointegration through balancing apatite-forming ability and osteogenic activity. Mater Sci Eng C Mater Biol Appl 2022:112647.
185. Liu H, Wang Y, Hou Y, Li Z. Fitness of chassis cells and metabolic pathways for l-cysteine overproduction in escherichia coli. J Agric Food Chem 2020;68:14928-37.
186. Liu H, Hou Y, Wang Y, Li Z. Enhancement of sulfur conversion rate in the production of l-cysteine by engineered escherichia coli. J Agric Food Chem 2020;68:250-7.
187. Yang D, Park SY, Park YS, Eun H, Lee SY. Metabolic engineering of escherichia coli for natural product biosynthesis. Trends Biotechnol 2020;38:745-65.
188. Choi KR, Jang WD, Yang D, et al. Systems metabolic engineering strategies: integrating systems and synthetic biology with metabolic engineering. Trends Biotechnol 2019;37:817-37.
189. Service RF. Synthetic biology. Synthetic biologists design ‘living materials’ that build themselves. Science 2014;343:1421.
190. Tang T, An B, Huang Y, et al. Materials design by synthetic biology. Nat Rev Mater 2021;6:332-50.
191. Depommier C, Everard A, Druart C, et al. Supplementation with akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat Med 2019;25:1096-103.
192. Riehl TE, Alvarado D, Ee X, et al. Lactobacillus rhamnosus GG protects the intestinal epithelium from radiation injury through release of lipoteichoic acid, macrophage activation and the migration of mesenchymal stem cells. Gut 2019;68:1003-13.
193. Ozdemir T, Fedorec AJH, Danino T, Barnes CP. Synthetic biology and engineered live biotherapeutics: toward increasing system complexity. Cell Syst 2018;7:5-16.
Cite This Article
Export citation file: BibTeX | RIS
OAE Style
Liu H, Geng Z, Su J. Engineered mammalian and bacterial extracellular vesicles as promising nanocarriers for targeted therapy . Extracell Vesicles Circ Nucleic Acids 2022;3:63-86. http://dx.doi.org/10.20517/evcna.2022.04
AMA Style
Liu H, Geng Z, Su J. Engineered mammalian and bacterial extracellular vesicles as promising nanocarriers for targeted therapy . Extracellular Vesicles and Circulating Nucleic Acids. 2022; 3(2): 63-86. http://dx.doi.org/10.20517/evcna.2022.04
Chicago/Turabian Style
Liu, Han, Zhen Geng, Jiacan Su. 2022. "Engineered mammalian and bacterial extracellular vesicles as promising nanocarriers for targeted therapy " Extracellular Vesicles and Circulating Nucleic Acids. 3, no.2: 63-86. http://dx.doi.org/10.20517/evcna.2022.04
ACS Style
Liu, H.; Geng Z.; Su J. Engineered mammalian and bacterial extracellular vesicles as promising nanocarriers for targeted therapy . Extracell. Vesicles. Circ. Nucleic. Acids. 2022, 3, 63-86. http://dx.doi.org/10.20517/evcna.2022.04
About This Article
Special Issue
Copyright

Data & Comments
Data



 Cite This Article 57 clicks
Cite This Article 57 clicks

 Like This Article 37
likes
Like This Article 37
likes







Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.